

了解旧石器时代人类的生存策略需要对动物遗骸进行全面的分类鉴定。晚更新世动物组合的高度碎片化常常妨碍基于骨形态的正确分类鉴定。人们一直认为,动物群中形态上不可识别的部分反映了形态上可识别部分的分类丰度。在这项研究中,我们利用胶原蛋白I型肽质量指纹图谱(ZooMS)分析了保加利亚Bacho Kiro洞穴和法国Les cott
和La Ferrassie洞穴中至上旧石器时代过渡(MUPT)的三个动物数据集。我们的研究结果强调,旧石器时代骨骼组合的碎片成分可能与形态上可识别的成分有很大不同。我们得到了不同分类群之间的识别率对比,结果表明在Les cott
和La Ferrassie,形态上已识别的驯鹿(Rangifer tarandus)的代表性过高,而野牛/野牛(Bos/ bison)和马/欧洲驴(Equus)的代表性不足。再加上动物组成相对多样性的增加,这些结果对尼安德特人和智人在欧洲可能相互作用期间的生存策略的解释具有启示意义。此外,动物群落组成和食肉动物活动的变化表明,在整个MUPT中,人类与食肉动物之间的相互作用发生了变化,并表明尼安德特人和智人在遗址使用上可能存在差异。传统和生物分子方法的结合使用允许(动物园)考古学家解决一些在旧石器时代骨骼组合的形态学评估中通常面临的方法限制。
在物质记录发生重大变化的阶段,对猎物选择和狩猎策略的行为变化进行调查,是理解人类行为进化、文化变异和种群动态之间关系的关键(Delagnes and Rendu 2011;Discamps et al. 2011;Niven et al. 2012;Rendu et al. 2012;斯蒂尔2004)。传统上,这种行为模式是通过分析从旧石器时代遗址出土的石器和动物遗骸来研究的。特别是,动物标本提供了识别和记录人类种群为开发其环境而发展的行为的机会(Gaudzinski和Roebroeks 2000;Gaudzinski-Windheuser et al. 2014;莫林2012;Pederzani et al. 2021;Smith等人2021;斯蒂娜1993)。然而,研究古代动物不仅提供了古环境信息,而且当与人类活动相关的骨骼表面变化分析相结合时,它可以微调人类职业的时间,并有助于重建人类的饮食和与其他群体甚至物种的互动(Steele 2015)。事实上,动物剥削与各种各样的行为和认知方面有关,这些行为和认知方面与流动性、社会组织、技术发展和生存能力交织在一起(marian和Assefa 1999)。
然而,晚更新世的骨骼组合通常是高度碎片化的,这使得仅根据形态学对许多标本进行正确的分类鉴定和解剖归属(Lyman 2002;Morin et al. 2017a)。有几个过程会影响动物遗骸,从分解、沉积物中的选择性破坏、死后运输和埋葬开始,以保存在挖掘、清洁处理和储存过程中可能发生变化的骨骼标本(Lyman 1994;马里恩1991)。所有这些因素,包括地质的、生物的和文化的,都可能导致区系鉴定的变异性。与差异保存一起,它们可以为分类群相对丰度的解释和量化创造潜在的偏见来源(Dirrigl 2002;马里恩1991;marian and Kim 1998;莫林2004;Morin等,2017a, 2017b;Pickering et al. 2006)。事实上,再加上人类和食肉动物活动的影响,这些因素导致了分类学上可诊断骨骼的比例减少,导致可识别标本的数量减少。这样的过程可能会严重扭曲各种考古和生态推断(Faith 2007;Morin et al. 2017a)。
生物分子方法的最新发展使我们能够利用保存在这些骨碎片中的胶原蛋白对动物标本进行分类鉴定(Buckley et al. 2009)。通过质谱(ZooMS)动物考古学的蛋白质组学筛选对高度碎片化的骨骼进行分析,用于旧石器时代动物组合的分类评估,已经证明了其巨大的潜力(Berto等人,2021;Brown et al. 2021b, a;Buckley et al. 2017;Pothier Bouchard et al. 2020;Ruebens et al. 2022;Sinet-Mathiot et al. 2019;Welker et al. 2015),并强调了在研究人类生存时使用多方法方法的必要性。因此,形态学鉴定和zooms鉴定组分的分类特性可以与骨表面修饰分析相关联,从而通过重建地形学历史来解决标本表面保存和骨积累因素。I型胶原蛋白的分析提供了基于氨基酸序列变化的分类识别,并允许骨组合的分类识别扩展到形态上不可识别的成分。以前应用zoom作为动物组合的筛选工具,在这两个组成部分的可比性方面提供了不同的结果。一个动物群中形态上不可识别的成分的分类丰度通常不会与形态上可识别的成分不同(Berto et al. 2021;Buckley et al. 2017;Welker et al. 2016, 2017),但这并不一定表明一种模式(Ruebens et al. 2022;Sinet-Mathiot et al. 2019)。此外,这种差异可能反映了与骨破碎和胴体加工强度相关的特定人类行为特征(sinet - mathio et al. 2019)。更好地了解变异的来源将有助于预测某些骨组合中可能发生的潜在差异。
动物考古学文献经常包含骨骼标本的体型分类属性,这些属性不能可靠地分配给特定的分类群或进化支。一般认为,这些体型分类属性是可靠的,反映或包含了整体骨骼组合的分类信息。然而,之前的ZooMS研究强调,这是一种潜在的不可靠方法(Sinet-Mathiot et al. 2019)。在这里,我们测试了三个晚更新世遗址的骨骼组合的碎片成分:Bacho Kiro洞穴(保加利亚)和Les cott
和La Ferrassie(法国)。它们都显示了丰富且保存完好的中-上旧石器时代过渡(MUPT)地层序列。这些遗址提供了讨论尼安德特人和晚更新世智人之间可能相互作用期间生存策略的历时变化的机会(Hajdinjak et al. 2021;highham et al. 2014;Hublin 2015;Hublin et al. 2020;普雷弗等人,2021)。这项工作探讨了将形态学上无法识别的骨骼成分分析纳入总体骨骼积累方面的动物群组合描述的含义,旨在推进我们对MUPT期间人类生存策略的解释。我们解决了在动物群形态评估过程中通常面临的方法限制,并展示了如何添加生物分子方法,如非靶向zoom筛选,可以通过提供更清晰的猎物选择和地点占领图像来补充我们对生存行为的理解。通过包括跨越欧洲MUPT的组合,我们因此能够证明,通过zoom对骨组合碎片成分的评估可以提供不同的物种频率模式,而不是以前仅基于形态学可识别的记录进行解释。
本研究包括对三个晚更新世遗址(Bacho Kiro Cave、Les cott
和La Ferrassie)的骨骼材料进行zoom分析;(图1,表1)。所有动物考古学家通过骨骼形态学进行分类鉴定的材料都被称为形态学成分。同样,所有通过zoom在形态学上无法识别和分类学上识别的碎片标本将被称为ZooMS组件。这三个地点都是最近发掘出来的,并且提供了大型的、环境良好的、高度碎片化的动物遗骸的骨骼组合。使用可比的动物考古学方法和协议对形态学鉴定和碎片性无法鉴定的骨骼组合进行骨表面分析。所有的区系数据均来自近期的挖掘活动,形态学和ZooMS组分的标本在挖掘区域显示出相似的空间分布。由于在遗址挖掘过程中,碎片大小的限制为20毫米,为了允许最终进行额外的生物分子分析,我们选择了长度通常大于20毫米的碎片和形态无法识别的碎片样本进行蛋白质组学分析。事实上,动物考古学对小于2厘米的碎片的分析通常受到限制,因为分类属性依赖于尺寸。考古遗址挖掘过程中泥沙筛分产生的骨材料不包括在本研究中。从La Ferrassie第6层动物群中选择所有形态上无法识别的标本进行ZooMS分析。在Bacho Kiro洞穴和Les cott
洞穴中,动物考古学家从形态学上无法识别的材料中随机选择标本,没有对骨骼形态进行任何选择,也没有对表面进行特定的修饰。
Bacho Kiro洞穴(保加利亚德里亚诺沃)位于巴尔干山脉(Stara Planina)的北坡,多瑙河以南约70公里处。以前在20世纪进行过调查(Garrod et al. 1939;Koz?owski和Ginter 1982),该遗址于2015年由国家考古研究所与保加利亚科学院博物馆(保加利亚索非亚)和马克斯普朗克进化人类学研究所(德国莱比锡)重新开放进行挖掘。考古序列跨越中旧石器时代(MP)到上旧石器时代(UP)。从第1层和第J层的两个区域(主要区域和生态位1)中发现的考古材料被认为是旧石器时代早期晚期的一部分,标志着晚更新世智人在欧洲最早出现(Hublin et al. 2020)。这从J层上部的45,990 cal BP开始,并在第1层显著增强,可追溯到45,040-43,280 cal BP (less et al. 2020;Pederzani et al. 2021)。该材料包含欧洲最早和数量最多的智人骨工具和装饰品组合,部分通过zoom进行分类鉴定(Martisius et al. 2022)。从技术上看,从K层恢复的组合与MP相关联,沉积于61±6000至51000年前(Fewlass等,2020;Pederzani et al. 2021)。我们通过zoom对生态位1和主要扇区的第1层(814个标本)、第J层(438个标本)和第K层(343个标本)的1595个动物遗骸进行了调查(Hublin et al. 2020)。根据先前描述的方法(Smith et al. 2021),对来自两个部门的I层、J层和K层的7,013具动物遗骸进行了动物考古学分析,包括分配给一个分类群的1,453个标本(I层1,077个,J层232个,K层143个)。
Les cottimac(法国维埃纳)是一个位于巴黎盆地和法国中西部普瓦图之间走廊的洞穴。该遗址于1878年被发现,并通过几次发掘活动进行了探索(Bastin et al. 1976;桑德琳。1997;Pradel 1967),但本研究中包含的材料来自于2006年由M. Soressi在法国文化和通讯部以及马克斯·普朗克进化人类学研究所的支持下开始的一次发掘(Soressi et al. 2010)。通过ZooMS,我们分析了523个形态上无法识别的动物标本,加上Welker et al.(2015)中提出的152个标本(137个无法诊断的碎片和15个在ZooMS盲测中分析的标本),总共有675个来自Les cottoms的标本被ZooMS分析。其中,220个来自Mousterian (US08,年代在46051 - 42034 cal BP之间,根据OSL测量在55 - 48 ka之间(Jacobs et al. 2015)), 217个来自ch
特佩龙期(US06,年代在42961 - 40344 cal BP之间),168个来自原Aurignacian (US04下),70个来自早Aurignacian (US04上)。在放射性碳年,奥震期地层的年代从40,372至36,697 cal BP (Talamo et al. 2012),或在OSL年从43至36 ka (Jacobs et al. 2015)。由于与其他层相比,US04上层的标本数量较少,因此需要谨慎考虑。骨表面分析是标准化的,以前在其他地方描述过(Rendu et al. 2019)。在通过传统动物考古学评估的总共5169块骨骸中,从亚科到种的范围内鉴定了1922块骨骼和牙齿标本(US08 397块,US06 166块,US04下部715块,US04上部629块)。
La Ferrassie的Grand Abri(法国Savignac-de-Miremont)位于法国西南部的Dordogne地区,位于vz
河的支流山谷中,在20世纪由Capitan和Peyrony首先挖掘,然后由Delporte (Delporte和Delibrias 1984;Peyrony 1934)。Turq及其同事在2010年至2015年进行的一次发掘进一步细化了跨MUPT的地层层序(gusamrin et al. 2015;Turq et al. 2012)。chalterperronian地层(第6层)的年代为45,100至39,520 cal BP (Talamo et al. 2020),标志着该地区最早出现了这种石器工业。在这一层中,形态学上可识别为分类单元的动物材料仅占骨骼组合的17.5%(142个标本),以驯鹿(Rangifer tarandus)为主。所有切片绘制的、形态不确定的标本都通过ZooMS处理(527个标本)。
本研究采用的变焦提取方案先前已部分描述(Buckley et al. 2009;van Doorn et al. 2011;Welker et al. 2016)。全部2645份标本(10 - 30mg)用钳子取样,置于96孔板中。可溶性胶原蛋白在100μL 50 mM碳酸氢铵(AmBic)缓冲液中65℃孵养1小时,提取可溶性胶原蛋白。为了改进和验证可溶性胶原蛋白的分类特性,440个标本(La Ferrassie 70个,Les cott
369个,Bacho Kiro Cave 1个)(SI表2)在130μL 0.6 M HCl中4℃脱盐18-20小时,中和至pH 7,再在AmBic中溶解。然后,用胰酶(0.5μg/μL, Promega)消化50μL所得上清液,在37°C下过夜,用三氟乙酸(20% TFA)酸化,然后在Hypersep C18 96孔板(Thermo Scientific)上用真空歧管清洗。简而言之,将96孔深孔板(Eppendorf)放置在Hypersep板下方以收集溶液。用200μL 0.1% TFA(50:50乙腈)和UHQ水(调理液)调理Hypersep板上的C18滤头,并用200μL 0.1% TFA和UHQ水(洗涤液)洗涤。肽提取物通过过滤器缓慢抽真空,以确保最佳的结合效率。所得废液被丢弃。然后用200μL洗涤液再次洗涤过滤器,并在100μL调理液中提取肽,转移到96孔板上。用多通道移液管(Thermo Fisher)在MALDI Bruker板上添加α-氰基-4-羟基肉桂酸(CHCA, Sigma)基质,标记三份消化后的肽段。
MALDI-TOF- ms分析是在德国莱比锡的Fraunhofer IZI进行的,使用自动旋转速度LRF MALDI-TOF (Bruker)在反射模式、正极性和高达590 Da的矩阵抑制下进行的,收集的质量电荷比范围为1000-3500 m/z。在R 4.0.5版本(R Core Team 2021)和MALDIquant v. 1.21版本(Gibb and Strimmer 2012)中,每个样本合并了三个副本。首先,我们使用移动平均线平滑强度,并使用TopHat方法去除基线。然后,对于每个样品,我们使用supersmother和信噪比为3对齐重复光谱,将三个重复相加得到单一光谱,并再次使用TopHat去除基线。光谱导出为。默沙东公司文件。采用mMass 5.5.0进行分类鉴定。(Strohalm et al. 2010)通过人工肽标记质量鉴定,与所有欧洲更新世大中型哺乳动物的肽标记系列数据库进行比较(Welker et al. 2016)。为了评估非内源性肽的任何潜在污染,我们在样品旁边进行了实验室空白。这些都是空的胶原肽,排除了现代实验室或储存污染的可能性。
在一些亲缘关系较近的物种中,肽标记序列可能是相似的,这是属于以下分类类群的物种的情况:Bos/Bison, Cervid/Saiga, Equidae和Ursidae。Cervid/Saiga可以归因于Cervus elaphus(马鹿),Megaloceros giganteus(巨型鹿),Alces Alces(麋鹿)或Dama sp.(黇鹿)。马科和熊科分别包括来自马属和熊科的物种,最有可能的是ferus和Equus hydruntinus或熊科和熊科熊科。为了便于对ZooMS和形态学组成进行比较,将最常见的物种和分类群分为Bos/Bison (Bos primigenius、Bison priscus和Bos/Bison sp.)、Cervid/Saiga (C. elaphus、D. dama、M. giganteus)、熊科(U. arctos、U. spelaeus和Ursus sp.)、Capra sp. (C. ibex和Capra sp.)和Equidae (E. ferus、E. hydruntinus和Equus sp.)(SI Table 3)。形态学成分的Cervid标本也被归入Cervid/Saiga类群。在ZooMS组分中,鉴定为Cervid/Saiga/Capreolus Capreolus的少数标本被纳入更广泛的分类类群Cervid/Saiga,以允许两者组分的可比性。
建议作为胶原保存的指标(Welker et al. 2017;Wilson et al. 2012),根据已发表的协议(van Doorn et al. 2012),计算所有样品中肽COL1α1 508-519的谷氨酰胺(Gln)脱酰胺率(Brown et al. 2021b, a),这在I型胶原的肽指纹图谱中经常观察到(van Doorn et al. 2012;Wilson et al. 2012)。脱酰胺率的范围从%Gln=1.0到%Gln=0.0表示谷氨酰胺完全脱酰胺。在常规ZooMS筛选过程中获得的谷氨酰胺脱酰胺率先前已被建议用于评估骨骼组合的同质性(地点内的空间和时间变异性),检测地层异常值(侵入性物质或差异骨保存),告知特定肽和标本的保存质量,或从生物分子角度查看分类分布(Sinet-Mathiot等人,2019;Welker et al. 2017;Wilson et al. 2012),尽管取得了不同程度的成功(Brown et al. 2021b, a)。
由各自的动物考古学家在形态学和zooms鉴定的标本上记录了所有的地貌学修改,并在研究地点内部和之间保持一致。形态学鉴定和未鉴定的骨表面均通过目视检查进行评估,必要时使用放大倍率(高达20倍放大倍率)(Blumenschine et al. 1996)。用数字卡尺分别测量骨标本的最大长度。
尽管在使用Stiner及其同事(Stiner和Kuhn 1995)提出的刻度(0:未燃烧到6:完全煅烧)的分类分析中记录了燃烧的痕迹,但由于胶原蛋白保存不良,这些烧焦的残留物被排除在随后的ZooMS分析之外。所有骨骼的风化阶段都被记录下来,并为理解埋葬前骨骼的暴露(短/长时间)提供了一个定性尺度(Behrensmeyer 1978)。对Les cott
骨骼组合使用了稍微修改的方案,记录了与风化有关的特定修改(见Rendu等人(2019))。具体来说,风化是根据三个变量来记录的:剥落(骨表面剥落)、开裂(骨表面出现纵向裂缝)和解体(骨的完全破坏)。此外,其他记录的修改包括牙根腐蚀和磨损(以受影响骨表面的百分比表示)。方案范围从0%(未观察到明显的改变)到100%(覆盖整个骨表面)(Smith et al. 2021;Behrensmeyer 1978;Blumenschine et al. 1996;Domínguez-Rodrigo et al. 2017;费舍尔1995;李·莱曼1994;Olsen and Shipman 1988;Soulier and Costamagno 2017)。
对于所有三种骨骼组合,人类的变化包括与屠宰和胴体加工有关的痕迹(切痕、刮痕、剁痕、骨髓断裂),食肉动物的变化包括牙印、啃咬痕迹、骨折和消化造成的损伤以及啮齿动物的牙齿痕迹。已鉴定标本数(NISP)是指分配给一个分类单元的标本数量。
当不可能在形态学上将骨碎片标本分配给特定的分类群时,根据先前的分配将其分配给特定的体型类(Morin 2012)。将特定分类群划分为不同体型的分类通常是在体型和骨骼大小的基础上进行的(Morin 2012;Rendu et al. 2019;Smith et al. 2021))。
ZooMS和形态学成分的结合可以评估骨骼元素的分布,并可能识别以前未被识别的骨骼元素,这对我们理解狩猎策略和尸体运输具有重要意义。由于骨骼元素是在分类学上无法识别的标本上识别出来的,我们的目标是将骨骼部分鉴定与ZooMS分类学鉴定联系起来,以评估形态学和ZooMS鉴定的成分之间的骨骼代表性。为了评估每种成分中优势分类群的骨骼成分代表,每种鉴定方法将骨骼成分分类为身体部位(颅骨:头盖骨,下颌骨;轴向:椎骨、骨盆、肋骨;前肢:肱骨、桡骨、尺骨;后肢:股骨、胫骨;远端肢体:腕骨、掌骨、跗骨、跖骨、指骨;LBN:长骨碎片,FBN:扁平骨碎片)(基于Stiner (1991a, 1991b))。在所有三个数据集中,牙齿和鹿角与颅体部分分开分类,因为它们的包含可能会使组件之间的比较产生偏差。事实上,鹿角和角芯往往是罕见的,更容易在形态学上识别,减少了它们在zoom成分中的代表性。解剖上无法识别的标本(NID)被排除在骨骼元素表征的评估之外,因为它们没有提供实质性的信息。
计算生态多样性指数,探讨添加zooms鉴定标本对各层、各立地动物群落分类多样性的影响。我们使用Shannon- wiener指数(H ') (Shannon 1948)和R package vegan v. 2.6-2 (Oksanen et al. 2019)来量化我们在zoom和形态学成分之间的三个动物组合的分类多样性,并评估每个站点层之间动物多样性的任何变化,同时考虑到分类丰富度及其丰度分布。由于Shannon-Wiener指数对样本量很敏感,所以样本量小时应谨慎考虑数值。Pielou均匀度(J′)与物种丰富度一起,通过给出每个分类类群在每个组分中的个体数量来衡量分类多样性,反映分类群间分布丰度的均匀性。取值范围为0(不均匀)~ 1(完全均匀)。
摘要
介绍
材料与方法
结果
讨论
结论
数据可用性
参考文献
致谢
作者信息
道德声明
补充信息
搜索
导航
#####
对所有三个数据集的ZooMS分析显示,保存完好的I型胶原具有很高的分类鉴定成功率,可达亚科或属的范围,在90%至97%之间(SI表1)。对于82%的样品,半破坏性提取方案(AmBic)足以获得ZooMS鉴定。在Bacho Kiro洞穴,胶原蛋白保存良好(Fewlass等人(2020)也注意到),导致判别性分类身份的比例很高。所有ZooMS样品仅使用AmBic协议提取,而我们通过酸脱矿提取了一个标本,以验证其分类身份(蓖麻纤维)。在Les cottoms和La Ferrassie,样品使用AmBic和酸脱矿方案进行处理,以改进和优化分类鉴定(SI表2)。
地层单位和分类群之间的脱酰胺作用
计算谷氨酰胺脱酰胺率是为了检测考古层之间的潜在侵入物质或分类群之间的胶原蛋白保存差异。由于数据非正态分布(Shapiro-Wilk正态检验,p值< 0.05),我们采用Wilcoxon-Mann-Whitney检验比较不同分类群和不同层间的谷氨酰胺脱酰胺率。这里给出的脱酰胺率是从使用无酸方案分析的样品中得出的。在Bacho Kiro洞穴,我们观察到来自K层的较老样本显示脱酰胺水平升高,且各层之间的值显著不同(SI表5,SI图2)。我们注意到,除了US06的脱酰胺值与US04和US08的脱酰胺值有显著差异(SI表5)外,Les cottoms各层之间的脱酰胺值重叠(SI图3)。优势分类群之间的谷氨酰胺脱酰胺率似乎重叠,这表明它们在每个位点内经历了相似的分子成岩过程(SI图4)。然而,可以确定一些例外。在Bacho Kiro Cave,各类群间的谷氨酰胺脱酰胺率具有相似性,特别是在第K层内,但在第I层和第J层,与其他类群的谷氨酰胺脱酰胺率存在显著差异,特别是与Bos/Bison、Capra sp.和Equidae相比(SI Table 6)。除了少数Rangifer tarandus标本(n=6)的脱酰胺值与US04 lower的Equidae有显著差异(wilcox - mann - whitney检验,统计值=64,p值=0.013,SI表6)。在La Ferrassie, Rangifer tarandus和Cervid/Saiga标本的脱酰胺值与Bos/Bison有显著差异(SI表6)。某些分类类群和层间的统计差异可能是样本量差异造成的。土语系的历史,和遗址形成或屠宰实践。然而,为了解释这些差异,需要进一步的探索。
在每个站点中,ZooMS和形态学成分之间的物种表征通常是一致的,但添加ZooMS可以识别通过形态学无法识别的分类群。在Les cottassis, ZooMS在US06获得的动物群落中鉴定出Felidae和Ursidae,但也导致在US04下层增加了Cervid/Saiga (SI表7)。在Bacho Kiro洞穴,ZooMS分析允许在J层鉴定象科(SI表8)。在La Ferrassie,使用ZooMS导致分类鉴定标本数量增加了四倍。因此,这一层的分类多样性扩大了,增加了Capra sp.、象科、Rhinocerotidae、Ursidae和一些食肉动物(SI Table 9)。
Shannon-Wiener指数计算表明,随着zoom的增加,一个站点上确定的动物群落的多样性会发生显著变化。更具体地说,我们观察到,在La Ferrassie、Les cott
和Bacho Kiro洞穴所研究的地层中,ZooMS和形态学组合的动物多样性有所增加(图1,SI表10)。相比之下,在Bacho Kiro洞穴,在添加ZooMS身份后,第一层动物组合的Shannon-Wiener指数值较低,表明分类多样性较低。这种模式可能强调了与更大样本量相关的形态学成分内更好的识别率,或者由于在形态学成分中显示低丰度的分类群的重复识别而突出了更高的ZooMS成分的均匀性。
图1
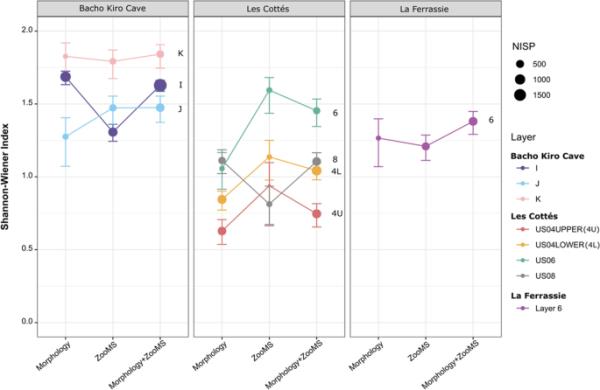
Bacho Kiro Cave、Les cott
和La Ferrassie洞穴各研究层的Shannon-Wiener指数比较了不同分类鉴定方法的差异(详见SI Table 10)。每个值给出置信区间(2.5-97.5%)
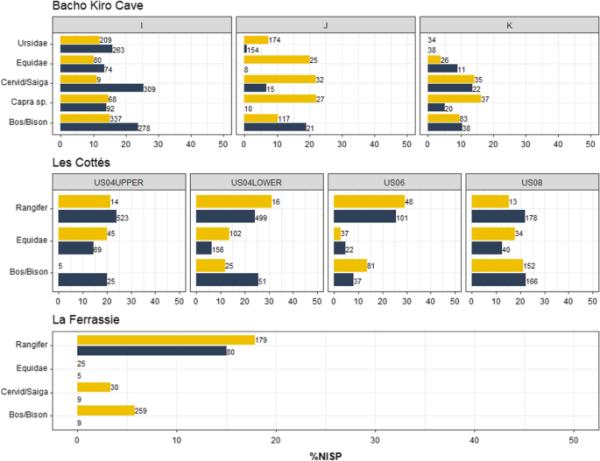
优势类群(即占比最高的类群)在各样点内的分布一致(Bacho Kiro Cave: Ursidae、Equidae、Cervid/Saiga、Capra sp.和Bos/Bison;羚羊:野牛、Equidae和Rangifer tarandus;La Ferrassie: Bos/Bison, Cervid/Saiga, Equidae和Rangifer tarandus),但我们观察到它们对整体骨骼组合的相对贡献存在差异(图2)。在Les cottims和La Ferrassie, zoom成分表明驯鹿的比例较低,被Bos/Bison和Equidae的较高比例所抵消(SI表7,9和11)。我们注意到La Ferrassie的野牛/野牛比例增加了9倍。在Les cottimass,我们观察到野牛/野牛和Equidae的平均数量增加了两倍,增加了zoom来分析动物群。在Bacho Kiro洞穴,与Les cottassie和La Ferrassie相似,在ZooMS组分中,Bos/Bison的残留物略多,特别是在I层和J层,相反,Ursidae的特征与La Ferrassie和Les cottassie的驯鹿相似,但在J层和k层的比例略低。我们注意到在I层中Cervid/Saiga的鉴定方法存在很大差异,但这些差异并不一致。在比较不同层间的区系组成以评估不同类群的NISP的变化或转移时,我们注意到,在Les cottacims,从US08到US04,野牛/野牛的比例逐渐减少,被Equidae的增加所抵消,这一点通过使用zoom尤其明显。尽管通过zoom分析的Les cottimacs早奥日尼亚世(US04上部)标本数量较少,但在优势分类群之间的分类丰度方面,所获得的结果与以下层的结果显示出连续的模式。
图2
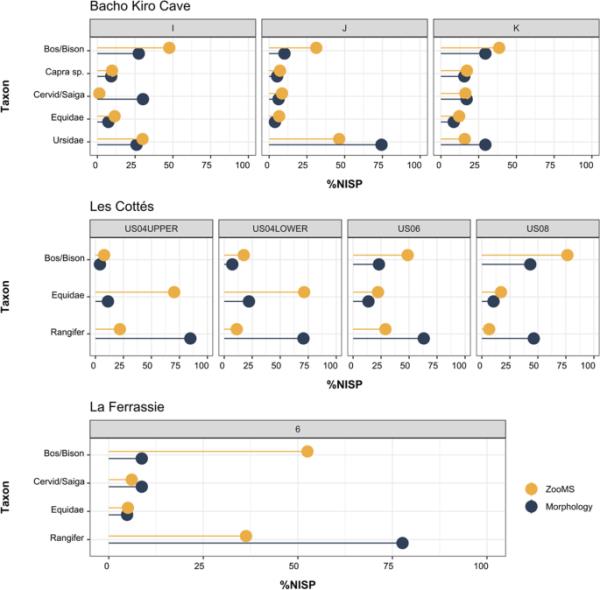
Bacho Kiro Cave、Les cott
和La Ferrassie的ZooMS和形态学组成的优势类群百分比
虽然在没有其他替代方法来解释该组合的这一组成部分时,将形态学上无法识别的标本分类为体型类别仍然是一个有用的工具,但ZooMS提供的分类鉴定与体型类别之间的相关性表明不一致。因此,之前在Fumane洞穴进行的观察似乎不是例外,而是常态(Martisius et al. 2020;Sinet-Mathiot et al. 2019)。我们观察到体型分类归属(主要基于骨骼大小和皮质厚度)与ZooMS分类归属之间的不一致(SI Table 4)。例如,熊科标本存在于大多数食肉动物和有蹄类动物的体型分类单位中(图3)。一些马科动物标本被归类为大型食肉动物类,Caprinae和Capra sp.被归类为大型有蹄类(图3)。尽管许多动物考古学家已经在使用替代命名法(即哺乳动物类别或未知类别,而不是有蹄类或食肉动物类别(Castel 2011))或标准化工具(Discamps 2021),但这些结果只是证实了身体大小类别归属应该谨慎使用。特别是在将这些分类转换为更具体的分类单位和/或评估人类生存策略时。当将骨标本分配给广义的科归属时,应谨慎避免“分类盲目性”,这种盲目性是基于皮质骨的厚度而假定的支系归属的丰富程度。
图3
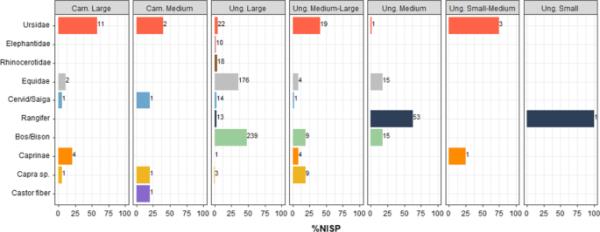
结合来自Bacho Kiro Cave, Les cott
正如预期的那样,较大的骨碎片通常更容易通过比较形态学来识别,因为它们通常保留更多的形态学特征。较小的碎片往往只能通过变焦来识别(图4)。这种模式在Bacho Kiro洞穴的第I层和第K层尤其明显(SI图5)。然而,并非所有分类群都是如此。我们注意到在优势分类群中,zoom和形态学成分之间的标本长度分布不同。在Bacho Kiro洞穴中,Bos/Bison、Cervid/Saiga和Equidae的标本长度呈相反的两极分布,而Capra sp.和Ursidae的标本长度分布较为相似。由于数据不是正态分布(Shapiro-Wilk正态检验,p值< 0.05),我们使用Wilcoxon-Mann-Whitney检验来比较不同分类群、层和鉴定方法之间的骨长分布。通过zoom鉴定为Capra sp.和Ursidae的骨标本,其片段长度分布与其他分类类群明显不同,特别是在第一层(SI Table 12)。同样,对Les cottims骨骼组合的观察表明,与驯鹿标本的相似分布和形态学成分分布相比,Bos/Bison和Equidae通常表现出相反的分布趋势(SI图6)。在Les cottims,形态学上鉴定为驯鹿的标本的骨长度分布与US04和US08的Bos/Bison和Equidae分布明显不同(SI表13)。但在ZooMS组件之间没有观察到差异。在比较不同鉴定方法的分布时,我们还注意到在US04和US08中野牛/野牛和马科动物标本的显著差异(SI表14)。由于没有对La Ferrassie的形态学鉴定成分进行公制测量,因此无法比较ZooMS和形态学成分之间的骨长度分布。然而,ZooMS组分占动物群组合的82.5%,因此ZooMS组分的优势分类群之间的标本长度比较是可能的。尽管来自优势分类群的标本通常表现出相似的长度分布,在2-3 cm范围内的比例很大,但马科动物的骨头往往具有较少的大碎片,在较小的尺寸类别内的标本比例较高(SI图7)。通过ZooMS识别的马科动物碎片的长度分布与Bos/Bison, Cervid/Saiga和驯鹿明显不同(SI表15)。最可能的原因是2-3厘米的马科动物碎片数量过多,而3-4厘米的标本数量不足。然而,应该注意的是,Equidae是样本量最小的分类群,这可能会影响这些结果。
图4
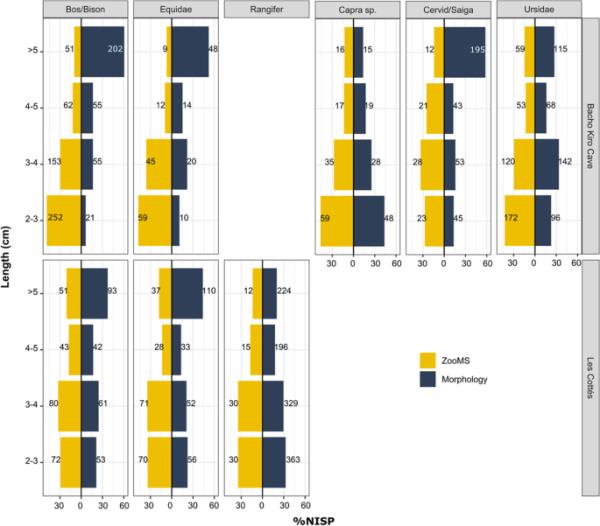
在Bacho Kiro Cave和Les cott
遗址中,优势类群在ZooMS内的骨长分布(橙色)和各研究层的形态组成(蓝色)。条形图上的数字是每个大小类别的NISP
骨表面保存
我们研究了骨表面的可读性,以排除与环境埋藏学因素相关的骨碎裂。我们发现,在Bacho Kiro Cave和La Ferrassie,通过zoom和形态学分类学鉴定的标本骨表面只受到低程度的表面风化的影响,这不能解释分类群和/或层之间破碎性的差异(SI表16和17)。
由于Les cottims的高度风化,ZooMS组件的许多骨骼表面呈现出自然断裂。特别是,来自US06和US08的大部分野牛/野牛碎片,以及来自US04的马科动物样本表明多种类型的表面损伤(SI图8)。这些模式在形态学成分中也记录在驯鹿身上,比例很高(US04和US06 > 50%)。表面的可读性反映了骨头是如何受到风化或其他可能导致碎裂的因素的影响的,与野牛和马科动物的骨头相比,驯鹿标本的可读性通常更好(SI图9)。
骨组合蓄能器
我们研究了与食肉动物和人类活动相关的骨修饰,以确定骨组合的蓄积剂。我们发现,在所有三个站点中,ZooMS分析允许改进分类身份与地学数据的关联,这在一些情况下提供了额外的行为信息。总体而言,在动物考古分析中包含ZooMS鉴定,突出了食肉动物和人类活动中表现出骨骼变化的各种分类群(SI图10和11)。这些结果对Bacho Kiro洞穴和La Ferrassie的J层和K层尤其有用,因为增加了三到四个以前与修饰剂(人类)无关的分类群。
在Bacho Kiro洞穴中,在Cervid/Saiga(第J层:3% NISP,第K层:23% NISP)和Capra sp(第J层:11% NISP,第K层:11% NISP)以及Bos/Bison (31% NISP)和Equidae (31% NISP)上,通过添加ZooMS,发现了食肉动物的变异。在第K层ZooMS组分中,食肉动物的变异影响了21%的优势分类群的遗迹,这比以前通过形态学获得的百分比要高得多(SI Fig. 12)。在La Ferrassie,与人类活动相比,第6层内食肉动物活动的比例相对较低,因为在ZooMS组件内仅在两个Bos/Bison标本上发现了食肉动物修饰(SI图12)。
除了食肉动物活动的证据外,在所有研究层内的大多数分类群上也存在人为改变。在Bacho Kiro洞穴J层的马科动物(20% NISP)和Capra sp. (22% NISP)身上发现了人类的修饰(图5),我们注意到J层Cervid/Saiga标本(22% NISP)上有相对较高比例的敲击痕迹(SI图11和SI图15)。在La Ferrassie,在Cervid/Saiga (4% NISP)和Bos/Bison (6% NISP)上发现了人类活动,但在马科动物标本上没有发现人类活动,在驯鹿遗骸上发现的敲击痕迹比例较高(9% NISP);图5和SI图13)。在Les cottimans,在所有研究层的优势分类群中,人类修饰的范围在10 - 20%之间,在驯鹿标本中(特别是US04和US06)发生的比例更高,主要表现为切割痕迹和敲击痕迹(图5和SI图14)。在Bacho Kiro洞穴和Les cott
,我们注意到从中旧石器时代晚期到旧石器时代晚期,食肉动物的活动逐渐减少,同时Bacho Kiro洞穴的人类活动增加,强化了之前描述的模式(Rendu等人,2019;Smith et al. 2021)。此外,我们注意到在Bacho Kiro洞穴的第1层和第J层中,来自不同分类群的食肉动物遗骸(n=93),如犬科动物(Canis lupus, Vulpes Vulpes),猫科动物(Panthera leo spelaea, Panthera pardus),洞穴鬣狗(Crocuta Crocuta spelaea)和熊科动物,反复出现人为修饰,而第K层只有两种食肉动物遗骸被人类修饰(SI Table 18)。在Les cott
,只有两个犬科动物标本显示出人类变异,而在La Ferrassie第6层的食肉动物遗骸上没有观察到人类变异(SI Table 18)。
图5
Bacho Kiro Cave、Les cott
和La Ferrassie遗址优势类群的zoom(橙色)和形态学(蓝色)组分的人为变化百分比。条形图上的数字是该分类群所鉴定标本的总NISP
由于其形态的特殊性,以及它们受地形学过程的不同影响,牙齿在形态成分中占很大比例,在骨骼元素中占最高比例,特别是来自Bacho Kiro Cave和Les cott
s的材料(图6和SI图16)。在Bacho Kiro洞穴,ZooMS成分的骨骼组成主要由长骨(LBN)、颅骨和轴向遗骸组成,ZooMS成分中轴向元素的比例较高,这可以解释为肋骨的过多代表(SI Table 19,图6)。肋骨元素很难分类识别,因为它们相对于骨骼的大小和比例没有保留许多特定的形态特征。长骨碎片(LBN)对应于来自前肢、后肢和远端肢体(掌骨和跖骨)的未知骨碎片。在ZooMS成分中被归类为LBN的骨标本主要由骨干碎片(来自中轴或骨骼骨骺附近)代表,但很少来自骨骺,如Bacho Kiro洞穴材料的例子所示(SI图17)。在Les cottoms的形态学组成中,我们观察到不同分类群之间的肢体残肢比例相对相似,除了在US06中没有记录到Bos/Bison的后肢和远肢残肢,但Bos/Bison和Equidae的颅骨标本比例更高(SI Fig. 16)。在La Ferrassie, ZooMS组件的元素表示只对形态学组件的骨骼表示有很小的贡献,因为大多数遗骸无法识别,也没有被分配到身体部位(SI Fig. 18)。
图6
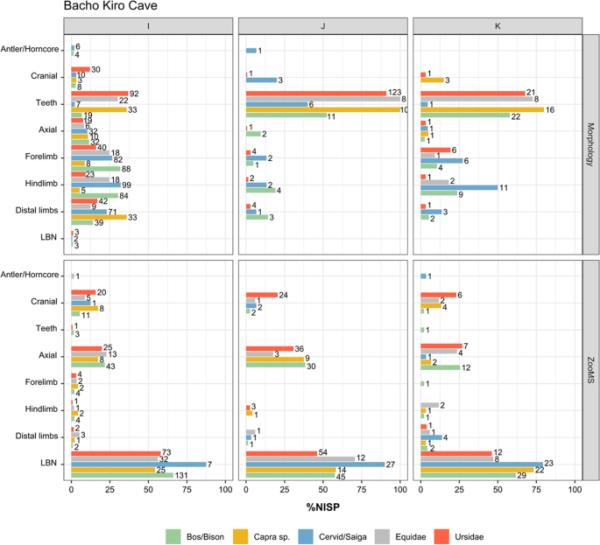
通过形态学(上)和zoom(下)鉴定的Bacho Kiro洞穴优势类群骨标本的骨骼分布。条形图上的数字给出了每个身体部位、层和ID方法的总NISP。未识别的身体部位(NID)被排除在情节之外。LBN,长骨碎片
这项研究结合了古蛋白质组学和动物考古学对三个数据集的动物材料的分析,这些数据集涵盖了中晚期旧石器时代的过渡。它的目的是克服由于骨头碎片导致的分类学鉴定的方法限制,并解决尼安德特人和晚更新世智人在欧洲可能相互作用期间的人类生存和动物加工行为。再加上分类学鉴定的高成功率,对骨骼组合中碎片化的、无法识别的成分进行zoom分析,可以识别以前通过传统形态学分析无法识别的物种,此外,还可以与传统的动物考古学、分类学和生态学数据进行整合和关联(Berto等人,2021;Sinet-Mathiot et al. 2019;Welker et al. 2015)。在高度碎片化的骨骼组合的情况下,这一补充可以为解释人类生存提供非常有价值的信息。这在我们对La Ferrassie layer 6的研究中得到了明显的例证,通过ZooMS进行分类鉴定的结果是形态学鉴定组分的4倍(NISPMorph=142, NISPZooMS=518)。
由于缺乏替代方法来解决旧石器时代骨骼组合的碎片化组成部分,先前对与生存策略相关的过去人类行为的研究仅依赖于形态学上可识别的动物,而排除了绝大多数可用的骨骼标本。然而,旧石器时代骨骼组合的碎片成分可能与形态可识别的成分有很大不同,突出表现在形态已识别的和ZooMS成分之间的优势分类群的比例差异。我们的研究没有反映在其他几个ZooMS筛选研究中观察到的模式,这些研究发现两个组分之间的优势种的分类组成相似(Berto et al. 2021;Buckley et al. 2017;Welker et al. 2016, 2017)。在这项研究中,这两个组成部分之间的分类丰度差异是通过在Les cottimass和La Ferrassie遗址中驯鹿的过度代表和野牛/野牛和马科动物的不足代表来观察到的。这些差异似乎与不同分类群之间的识别率差异有关,可能会根据其识别难易程度在优势分类群的代表中产生报告偏差。因此,驯鹿或熊科等分类群即使在碎片化的情况下也很容易区分,因此在形态学鉴定的成分中会被过多地代表。相反,当碎片化时,野牛/野牛和Equidae更难区分,通常被归类为无法识别的遗骸。
Bacho Kiro Cave和La Ferrassie遗址骨材料低风化模式的一致性表明,在整个地层中,自然因素对骨破碎的作用有限。总的来说,在这两个地点,骨材料被掩埋得相对较快,再次暴露的可能性最小。在Les cottims,虽然驯鹿的骨骼表面可读性稍好一些,但其风化程度在优势类群中是相当的。需要进一步的研究来了解这种模式是否可以通过骨骼形态或驯鹿标本的特定沉积条件(埋葬前较短的标本暴露)来解释,特别是知道谷氨酰胺脱酰胺率并不能表明明确的分子保存差异。此外,我们对每个位点的胶原脱酰胺分析并不能为每个组合的两个骨组分之间的分类比例差异提供分子成岩解释。当将ZooMS鉴定纳入动物考古学分析时,我们应该记住,根据定义,这两个组成部分通常显示不同的骨长度分布,因为较大的碎片往往更容易在形态学上识别。然而,在比较分类群时,我们注意到并非所有分类群都是如此(Pickering et al. 2006)。某些分类群,如Les cottacimos的驯鹿和Bacho Kiro Cave的Capra sp.,可以显示出与其他分类群(Bos/Bison和Equidae)明显不同的骨长度分布,这可能是由于在骨髓提取过程中最可能产生的骨碎片的大小和这些分类群之间不同的识别率。事实上,由于相对于骨直径的皮质厚度较低,并且与野牛和马科动物相比,它们的体型较小,驯鹿碎片将按比例覆盖更多的驯鹿骨头,这将使它更有可能保留可识别的特征。另一方面,掰开大型动物的骨头,如野牛或马科动物,平均会产生更大的碎片。牛骨标本的碎片通常很难与马科或马鹿的同源部分区分开来,因为这些分类群的骨骼元素往往在大小和形态上重叠(Morin 2012)。然而,由于驯鹿更容易识别,这导致该物种在形态学成分中的代表性增加,同时已识别的野牛/野牛和马科动物标本的比例有限(Gobalet 2001)。
猎物骨骼部分分布的评估往往与骨骼标本的分类鉴定密切相关。由于骨骨干缺乏诊断特征,加之骨髓提取导致的高碎裂率,小的长骨轴碎片往往难以识别(Morin et al. 2017a)。因此,通过zoom分析的形态学上无法识别的成分通常包含高比例的长骨,特别是骨干部分和肋骨碎片,对骨骼分布的评估提出了挑战。骨骺部分倾向于保留更具体的形态标准,便于遗骸的分类鉴定。然而,它们在ZooMS成分的长骨部分中的表现与Bacho Kiro洞穴的形态成分没有很大的不同。因此,骨骺代表性不足也可能是由于各种因素造成的选择性破坏,如不同的保存和骨密度,食肉动物的活动,特定的屠宰做法,如提取骨脂,以及考古挖掘过程中的沉积后或采样偏差(Binford 1981;格雷森和德尔佩奇2008;2010年、2020年;Yravedra and Domínguez-Rodrigo 2009)。诸如胴体加工和不同身体部位的选择运输等行为推断通常基于骨骼部分的代表性和丰度(Bartram等人,1999;Binford 1981;Klein et al. 1999;marian and Assefa 1999)。将骨骼表征与通过ZooMS获得的分类鉴定相结合,有可能为动物记录的清单增加元素,有助于我们理解铰接遗骸的运输。
然而,进一步将ZooMS数据与标准动物考古调查相结合,将需要两个组件中使用的度量具有更好的可比性。由于最小骨骼元素数(MNE)和最小个体数(MNI)不能与ZooMS数据进行定量比较,因此通常使用已识别标本数(NISP)(Grayson 1984)作为ZooMS组分中物种丰度的代表。如果骨碎片是基于它们的分类识别,而不考虑形态学或表面保存,可以建立实验模型,这将有助于我们更好地了解旧石器时代动物组合中的骨碎片如何根据标本大小而变化,以及识别骨骼元素。变焦可以帮助改进计算NME的替代方法,如骨元素的诊断地标或改装骨轴(marian et al. 2001;Morin et al. 2016;斯蒂娜1994)。
通过肽质量指纹图谱,将分类学鉴定的骨骼标本添加到跨越人类进化过渡阶段的动物组合中,有助于我们理解在MUPT期间观察到的转变模式。我们的研究结果进一步详细说明了在这一时期,古人类的饮食由一系列中型和大型食草动物主导(Discamps et al. 2011;Gaudzinski-Windheuser and Niven 2009;Gaudzinski-Windheuser and Roebroeks 2011;Jaouen et al. 2019;Niven et al. 2012;Rendu et al. 2019;Richards et al. 2008;史密斯2015年)。我们的工作强调了人类和食肉动物对更多样化分类群的开发,允许某些分类群与特定因素的相关性,这些因素有助于现场的骨骼积累,特别是在Bacho Kiro洞穴。在优势类群中,人类修饰主要由切割痕迹组成,骨髓提取的敲击痕迹发生率较低,因此无法解释成分之间比例的差异。ZooMS分析强调并细化了Les cottimac地区整个地层中分类群比例的变化,特别是马科动物和野牛/野牛标本之间的变化(Rendu et al. 2019)。这些动物组成的变化可能代表了遗址周围环境中猎物可用性的缓慢变化,或者与晚更新世智人在欧洲的扩张相似的人类选择策略。尽管如此,虽然形态学上确定的动物群表明更专注于狩猎驯鹿(Rendu et al. 2019),但我们的研究结果表明,这低估了对其他物种,特别是Equidae的开发。在关于驯鹿狩猎专业化的辩论框架内,这些结果特别令人感兴趣(Grayson and Delpech 2002;梅拉斯2004年)。尽管驯鹿在MUPT过渡期间的逐渐增加与MIS3期间的逐步气候退化相关,并且可以通过人类群体对环境波动的适应来解释(Banks等人,2013;Discamps et al. 2011),在整个MUPT中,大型有蹄类动物在人类饮食中的作用可能由于识别率的差异而被低估。
在尼安德特人占领这些遗址的后期,食肉动物变异的发生率表明,人类和食肉动物在动物的积累和变异中都很重要,这仍然表明人类经常占领洞穴,食肉动物偶尔来访,但后者可能比以前认为的更频繁(Straus 1982)。在Bacho Kiro Cave和Les cottimass,从MP到UP的食肉动物变异的减少突出了食肉动物活动的逐渐减少,这与之前在欧洲其他一些地点从这一时期发现的模式相吻合(Discamps 2014;Discamps等人,2019;Rendu et al. 2019;Smith等人2021;Stiner and Kuhn 2006)。在Bacho Kiro洞穴的IUP期间,在Ursidae遗骸上出现的人类修饰(以及在Bacho Kiro洞穴对其他食肉动物物种的修饰(Smith et al. 2021))强调了食肉动物与人类从竞争对手到猎物或原料来源的关系可能发生的变化。智人开始更密集地利用食肉动物遗骸作为原材料,特别是在IUP期间,在Bacho Kiro洞穴和东南欧和西南亚的其他地点,由洞穴熊骨头和牙齿制成的骨制品的增加(Bosch等人,n.d;Guadelli et al. 2011;Kuhn et al. 2009;Martisius et al. 2022;Stiner et al. 2013)。可以通过骨骼部分表示和胴体加工来调查原材料的这种特定需求(Rendu etal . 2019)。此外,在Bacho Kiro洞穴和Les cottimacs旧石器时代中期地层中较高比例的食肉动物痕迹证明,它们对该遗址的重复使用可能与人类占领的时间较短有关(Hublin et al. 2020;Smith et al. 2021)。在MUPT期间,人类群体与大型食肉动物之间的相互作用似乎发生了变化,这可能表明人类群体对其环境的掠食压力越来越大(Stiner and Kuhn 2006),或者与晚更新世智人相比,尼安德特人占据遗址的时间更短。
在对分类学上无法识别的标本进行分类分析的同时,应系统地对旧石器时代骨骼组合的碎片成分进行放大筛选(例如,参见Discamps(2021))。此外,将从考古遗址沉积物中提取的aDNA获得的动物群数据与动物考古学和旧石器时代动物群组合的ZooMS分析相结合,有可能更好地了解遗址的各种占领事件,或告知沉积物中保存的DNA的潜在起源。
下载原文档:https://link.springer.com/content/pdf/10.1007/s12520-023-01830-4.pdf

