

僧众猴(僧众猴属)是一种食果灵长类动物,它们喜欢吃种子,以树叶和昆虫来完成它们的饮食。众所周知,果肉和种子在成熟过程中具有不同的营养特性。种子的消耗可能是对资源可用性变化的一种适应,因为未成熟的种子比成熟的果肉或幼叶更稳定地可用。本文首次对僧猴(Pithecia monachus)的摄食生态学进行了研究。我们调查了秘鲁亚马逊地区área de Conservación区域社区Tamshiyacu-Tahuayo内季节性洪水森林的膳食组成,并确定了重要的取食植物。在20个月的时间里,我们徒步或乘独木舟跟踪了一群僧面猴,记录了459次进食事件。种子是最常食用的食物(49%),其次是果肉(中果皮、果皮或假种皮);25%)和节肢动物(22%)。树叶、树皮和花只是偶尔被摄入。成熟种子和节肢动物在僧面猴饮食中的重要性与其他研究不同:我们记录了大部分成熟种子的消耗,节肢动物的份额相对较高。
在热带森林中,水果是许多脊椎动物的重要食物资源(Fleming et al. 1978)。果实含有几种营养丰富的成分,这些成分具有不同的特征,随着发育阶段的变化而变化,并且在不同的分类群之间可能存在差异(Janzen 1983)。果肉可以由含糖的果皮或中果皮或富含脂质的假皮形成(Janzen 1983;Norconk 2021)。种子还含有高比例的蛋白质,但通常是物理和化学保护,以防止种子捕食(Janzen 1976;Norconk and Veres 2011)。一种特别依赖水果作为资源的脊椎动物是灵长类动物(Richard 1985)。所有南美灵长类动物(Platyrrhini)都依赖水果作为其饮食的一部分,尽管饮食中的比例和水果的哪一部分被使用差异很大(Rosenberger 2020)。
由于sakis很难适应并且非常害羞,因此对Pithecia sp.的摄食生态学研究不足(Pinto et al. 2013)。在Marsh(2014)研究的16个物种中,只有8个物种的摄食生态信息是可用的,而且往往局限于较短的研究时间或很少的观察。大部分研究都是在贵南地区进行的。只有两项研究对季节性洪水栖息地的sakis进行了研究,即P. rylandi脚注1 (Palminteri et al. 2012)和P. isabelaf脚注2 (Soini 1987)。
Sakis是食果动物,对种子有强烈的偏好,在以前的研究中,种子占其饮食的53-70% (Norconk和Conklin-Brittain 2004: 63%, P. pithecia;Palminteri et al. 2012: 70%, P. rylandsi;Peres 1993: 53%,白色假单胞菌)。种子通常被咀嚼,因此sakis充当“种子捕食者”(Ledogar et al. 2013;Norconk 2021)。它们具有高度专业化的牙齿形态,用于在吞咽前敲开硬壳水果和咀嚼种子(Kay et al. 2013;Kinzey and Norconk 1990;Norconk and Veres 2011)。种子的消耗被描述为对果实可用性变化的适应,因为它们是比成熟果肉更稳定的可用资源(Norconk 1996;Palminteri et al. 2012)。和果肉一样,种子在果实成熟的过程中会改变其化学成分(Norconk and Conklin-Brittain 2004)。例如,委内瑞拉sakis食用的某些植物在种子成熟期间,脂质水平上升,单宁水平下降(Kinzey和Norconk 1993)。先前的研究作者发现,各种sakaki物种消耗的种子中> 95%是未成熟的(Norconk 1996: 100%, P. pithecia;Oliveira et al. 1985: 100%, P. chrysocephala; Palminteri et al. 2012: > 99%, P. rylandsi;Peres 1993: > 98%,白色假单胞菌)。
Sakis以果肉、树叶和昆虫作为补充(Happel 1982;Izawa 1975;Kinzey 1992;Ledogar et al. 2013;Norconk 1996;Norconk and Setz 2013;Oliveira et al. 1985;佩雷斯1993;索伊尼1987)。在食用果肉时,sakis更喜欢成熟的中果皮和果皮(Charpentier et al. 2015;Norconk 1996;佩雷斯1993)。节肢动物并不总是被列为饮食的一部分,或者只占摄入食物的10%以下(Cunningham and Janson 2006: < 10%, P. pithecia;Kinzey and Norconk 1993: < 6%, P. picthecia;Peres 1993: < 1%,白色假单胞菌)。然而,食用昆虫可以在营养上补充果性饮食(Rothman et al. 2014;Urbani et al. 2019)。
Sakis分布在整个亚马逊流域,栖息在不同类型的森林中,包括高地原生林和季节性淹没林,白水várzea和黑水igapó (Marsh et al. 2018;Palminteri and Peres 2012)。前者通常由于每年洪水期间的营养沉积而提高生产力(Junk 1997;Melack和Forsberg 2001),尽管这在亚马逊西部地区不太明显(Prance 1979)。Sakis使用多种植物分类群,包括许多在其他platyrrhine物种的饮食中不那么重要的植物分类群(Boyle等人,2016;Norconk 2021)。Moraceae, Fabaceae, Chrysobalanceae, sapotacae, Annonaceae和Lecythidaceae等植物家族已被反复报道为其饮食的一部分(Charpentier et al. 2015;为了给1982;Norconk 1996;佩雷斯1993;Setz 1993)。Pitheciine生物量与Eschweilera trees (Lecytidaceae)的丰度呈正相关(Stevenson 2001)。Inga(豆科)、Brosimum (Moraceae)和Pouteria (saptacae)在不同栖息地的鸡类饮食中尤为重要(Boyle等人,2016)。
本文首次对僧面猴(P. monachus)的摄食生态学进行了研究。我们确定了僧面猴的饮食组成,并确定了重要的取食植物。根据以往对僧面猴取食生态学的研究,我们预计未成熟的种子是僧面猴食用最多的食物,其次是成熟的果肉。我们预计节肢动物偶尔会被吃掉,但不会在饮食中占很大比例。由于Eschweilera, Pouteria和Inga在淹水森林中存在,并且据报道是其他saki物种的重要食物,我们希望这些也包括在我们研究地点的僧侣saki饮食中。
área de Conservación地区社区Tamshiyacu-Tahuayo (ACRCTT)位于秘鲁亚马逊北部的洛雷托省。它最初成立于1991年,是一个保留地社区,但在2009年得到了额外的保护,并扩大到目前的42万公顷(Gobierno Regional de Loreto 2009)。ACRCTT以其高生物多样性而闻名,是13种灵长类动物的家园(Heymann and Aquino 1994;Valqui 2001)。在被指定为保护区之前,包括僧面猴在内的灵长类动物在该地区被猎杀(Bodmer 1995;newwing and Bodmer 2003)。该地区主要由未被洪水淹没的陆地栖息地组成,但也包含季节性洪水淹没的森林(Gobierno Regional de Loreto 2009)。塔华约河流域月平均气温为25 ~ 27℃,年降雨量约为3000毫米(Myster 2015)。
我们收集了亚马逊研究中心(ARC)周围的数据(4°22 ' 23”-4°24 ' 16”S 73°14 ' 45”-73°16 ' 36”W,图1)。ARC位于塔瓦约河的洪泛区,是亚马逊河的一条支流,由生态旅游公司亚马逊探险公司(www.perujungle.com)运营。塔瓦约河主要是一条酸性的黑水河流,营养含量低(Myster 2015)。河水水位在11月中旬左右上升,下层生态系统在1月底开始泛滥,从3月下旬到5月底(“汛期”)经历了大范围的洪水(Gobierno Regional de Loreto 2009;Ronchail et al. 2018)。从6月到10月,河流水位较低(“旱季”)(Gobierno Regional de Loreto 2009)。当提到我们研究区域的“旱季”时,我们指的是降雨较少、水位较低的月份,尽管全年降水量都很高(Kelly et al. 2014)。
图1
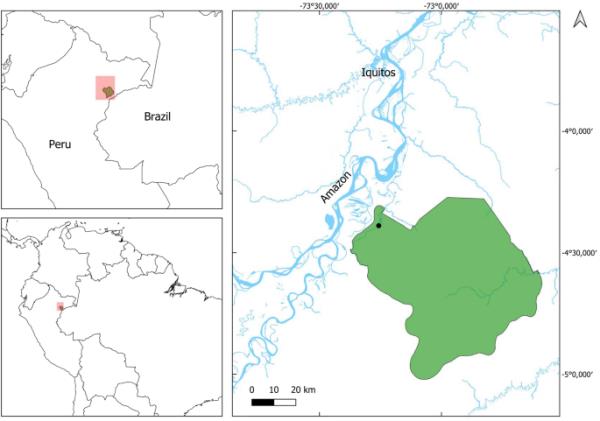
显示亚马逊研究中心位置的地图(ARC;●)在área de Conservación区域社区Tamshiyacu-Tahuayo (ACRCTT;绿色多边形)。地图右下角的河流(Rio Yavarí)标志着与巴西的边界
我们的研究物种僧猴(P. monachus)分布在秘鲁和巴西,位于亚马逊河/Solim?es、乌卡亚利河中下游和juru
河下游之间的流间区(Marsh et al. 2018)。它们普遍的害羞行为和不显眼的颜色对进行monachus的实地观察构成了重大挑战(Bartecki和Heymann 1987;Pinto et al. 2013)。除了作为各种捕食者的猎物之外,这些猴子由于其在该地区被猎杀的历史而保持神秘(Marsh 2014)。
我们在ACRCTT及其缓冲区内跟踪了至少12组僧面猴。群体大小在2到7个人之间变化,大多数群体由5个人组成(中位数=平均值=5)。每个群体至少由一个成年雄性和一个成年雌性组成(很容易通过它们的性别二色区分),有些群体有两个以上的成年个体。青少年分为11组。在研究中,婴儿分为两组。
我们从2019年7月到2020年7月,从2021年8月到2022年5月,在ACRCTT中搜索并跟踪了sakaki猴群体。saki组不习惯,通常非常害羞(Jackson 2016;Lehtonen 2017;Stenzel 2017)。我们在清晨5点到7点之间离开了ARC。我们轮流搜索下游(ARC以北)、上游(西南)和远离河流的内陆(东南)。我们在旱季花费了931小时寻找sakis,在汛期花费了1463小时(表1)。在旱季,我们使用独木舟向上游或下游移动,然后在找到一组sakis后步行跟随。在旱季,内陆的搜寻工作是徒步完成的。在汛期,我们用小船在河上移动,找到一群人后就换成了独木舟。在洪水季节用独木舟在内陆搜寻。我们要么通过视觉,通常是通过观察它们的运动,要么通过听到它们的叫声来定位它们。一旦找到一群人,我们就尽可能长时间地跟踪他们。我们将接触时间定义为我们与sakis进行视觉接触的时间,或者我们知道sakis在哪里的时间,例如,藏在树上。在后一种情况下,我们不断地观察隐藏的地方,以发现运动或观察喂食。如果可能的话,我们跟着一群日本人一直到天黑,然后在第二天早上日出之前回来。与每组的总接触时间在4到90小时之间变化(总努力=614接触小时)。
表1花费在搜索sakis上的时间,包括co每个月的接触时间和观察喂养事件研究期第n期(2019年7月至2020年7月、2021年8月至2022年5月)
我们使用行为抽样(Martin and Bateson 2007)在双筒望远镜的帮助下收集进食数据。当我们观察喂食时,我们记录了日期、时间和sakis的数量。我们将进食事件定义为一个个体对特定食物的进食,与他们进食的时间或摄入的食物量无关。例如,如果我们看到一群5只sakis在吃一棵树上的种子,我们记录5次进食事件,而不管每只sakis吃了多少种子。因此,如果我们看到一只崎在同一枝上吃几只蚂蚁,我们就记录了一次进食事件。如果saki以植物部分为食,我们记录下该植物的全球定位系统(GPS)位置,用胶带标记以备将来识别,并收集果实、叶子和树皮样本,供伊基托斯Amazonía秘鲁国立大学AMAZ植物标本室的植物学家鉴定。
我们将食用的食物类型分为种子、中果皮、果皮、假种皮、叶、树皮、花或节肢动物。对于水果部分的每次喂食事件,我们分配了成熟度类别:成熟,未成熟,成熟+未成熟(如果在同一喂食事件中食用成熟和未成熟的水果),干燥和未成熟(如果我们无法分配类别)。我们收集了sakis掉落的水果,检查哪一部分被消耗了,并与地面上完整的水果进行了比较。我们将果实储存在70%的乙醇中,并用我们分配给各自喂养树的编号标记小瓶。我们通过观察果实的大小和颜色来初步确定成熟度,并通过打开大小相近的完整果实来评估种子和果肉发育的阶段。如果可能的话,我们拍摄了取食植物和事件的照片(图2)。在我们的实地研究即将结束时,我们通过将储存的水果样本与全年收集的成熟水果进行比较,纠正了一些我们对成熟度的分配。如果果肉被食用,我们根据Cornejo和Janovec(2010)和van Roosmalen(1985)对果肉的属分类,指定果实的哪一部分变成肉质并构成果肉(中果皮、果皮或假种皮)。对于节肢动物,我们记录了从茎、枝、叶、附生植物或空气中采摘物品的基质。如果看得见的话,我们就记下被吃掉的节肢动物的纲、目或科;不可能有更高的分类学分辨率。
图2

僧猴正在吃红掌(天南星科)(图片来源:M. Gottstein)
摘要
介绍
材料与方法
结果
讨论
数据可用性
笔记
参考文献
致谢
作者信息
搜索
导航
#####
种子是最常食用的食物(49%,n=226),其次是节肢动物(22%,n=99)、中果皮或果皮(14%,n=64)、中果皮或果皮加种子(7%,n=33)、假种皮加种子(4%,n=20)、叶子(2%,n=10)、树皮(1%,n=5)和花(< 1%,n=2)。大多数对果实(种子、中果皮、果皮和假种皮)的取食来自成熟的果实(表2)。
表2果实(种子、中果皮、果皮、假种皮)摄食事件数及其成熟度
研究结果表明,猕猴取食植物212种,其中属级植物108种,种级植物72种。食用植物来自29个植物科(表2)。sakis食用的植物种类包括乔木(n=49)、藤蔓(n=14)、灌木(n=5)、棕榈(n=1)和附生植物(n=1)。大多数取食事件涉及Eschweilera (n=53,卵黄科)和Pouteria (n=30,仙人掌科)植物。我们无法确定消耗的干种子、树皮或花的种类(表3)。
表3植物种类及项目数在2019年7月至2020年7月和2021年8月至2022年5月期间由僧徒饮用
sakis以从树枝(n=27)、叶子(n=24)、附生植物(n=11)、空气(n=5)或树的主干(n=3)上捡来的节肢动物为食。在29个节肢动物摄食事件中,我们没有看到被摄食节肢动物的基质。我们观察到白蚁(n=5)、蚂蚁(n=4)、蝈蝈(n=4)、小蜘蛛(n=2)和蝴蝶(n=1)的食用量。我们无法对剩余的大多数节肢动物进行分类(n=83)。
观察sakis通常是困难的,在我们的研究地点遇到的群体都很害羞。我们在田野里呆了许多天,都没有见到sakis。特别是在旱季,观察进食非常困难,因为我们在森林地面上行走时会发出声音,而sakis在发现我们之后的一天里都会躲起来。因此,观测次数在某些月份相对较少。尽管如此,我们还是能够提供关于崎食性生态学的知识,这可以帮助我们发现物种和栖息地的差异。
高比例的种子和果肉分别在饮食中支持僧面猴分类为以种子为重点的食果动物。然而,我们只能部分地证实我们的预期,即未成熟的种子将是僧面猴最重要的食物来源,因为大多数食用的种子已经成熟。我们可以肯定,成熟的果肉将是僧袍酒的第二重要资源。
成熟的种子在营养上可能与未成熟的种子不同。例如,委内瑞拉sakis食用的成熟种子比未成熟的种子含有更高的脂质和游离单糖,而粗蛋白质则更低(Norconk and Conklin-Brittain 2004)。在不同季节收集数据的其他研究发现,饮食组成发生了变化,例如成熟水果的比例发生了变化,增加了对某些植物物种的使用,或消耗了节肢动物和树叶(Kinzey和Norconk 1993;Norconk 1996;Palminteri et al. 2012;索伊尼1987)。这些变化似乎不能在不同的栖息地和地区推广。我们观察到大多数摄食事件(86%)发生在汛期。虽然我们没有进行物候调查,但成熟和未成熟种子的可用性可能在一年中发生变化,这对sakaki的摄食生态有影响。由于在干湿季节的观察次数不同,我们没有研究饮食组成和饲料植物使用的可能的季节性模式。然而,日粮中成熟种子的高比例可能是由于资源可用性的季节变化。
与我们的预期相反,我们发现节肢动物经常被吃掉,占进食事件的第三高(22%)。这种差异与其他研究的一个可能解释是,我们使用了不同的量化喂食观察的方法。虽然我们使用喂食事件的数量来衡量一种资源在崎鱼饮食中的重要性,但大多数其他研究使用喂食时间作为衡量标准(例如,Cunningham和Janson 2006;Kinzey and Norconk 1993;佩雷斯1993)。由于节肢动物的消耗可能比打开果实以种子为食所需的时间要短,因此,当以事件而不是进食时间来确定节肢动物在sakis饮食中的相对重要性时,节肢动物在sakis饮食中的相对重要性可能要高得多。然而,我们清楚地表明节肢动物是僧面猴的重要资源。另一个可能的解释是我们研究发生的时间和栖息地。在季节性洪水泛滥的森林中,节肢动物在洪水季节可能更容易为sakis提供,因为一些分类群在洪水期间迁移到较高的森林地层中(Adis 1992;·伊勒姆1979;Souza et al. 2020)。从崎崎粪便中研究节肢动物多样性的作者认为,由于在野外观察崎崎的困难,节肢动物在其饮食中的重要性可能被低估了(Jesus et al. 2022;Pickett et al. 2012)。
共鉴定出29科70种取食植物。利用最多的植物科是卵磷脂科,其中消耗最多的是来自Eschweilera的种子。Eschweilera已被证明对sakis特别重要(Stevenson 2001),据报道,在亚马逊西部的季节性洪水森林中,P. isabela最常食用其种子(Soini 1987)。Palminteri等人(2012年)也描述了在某些月份增加使用毛柔沙的情况。在meta研究(Boyle et al. 2016)中,并非所有被发现对果蝇具有特殊重要性的植物属都被记录在我们的观察中:Pouteria的种子和果皮是第二大食用食物,但因加(Inga)的重要性较低(n=5),溴铵(Brosimum)根本没有记录。此外,在以前的研究中被报道为重要的桑科和番荔枝科植物在我们的观察中没有出现。然而,我们并不能识别出sakis所吃的所有植物。在我们的研究地点,植物的高度多样性和超年的结果模式使得我们可能只记录了僧面猴实际使用的植物的一小部分。
总之,我们在亚马逊西部季节性洪水泛滥的森林中发现僧面猴与其他不同栖息地的僧面猴有着相似的摄食生态。然而,我们也发现了一些差异。种子在成熟状态下被大量消耗,节肢动物的作用比文献中预期的更为重要。和尚可能略有不同的萨基斯喂养策略比其他日本米酒物种,虽然我们怀疑这些饮食差异主要源自于环境条件。这将有助于研究邻近的僧面猴种群,这些种群被限制在非洪水森林中。需要进行研究,以更好地了解sakaki的饲养生态,特别是在亚马逊西部。
下载原文档:https://link.springer.com/content/pdf/10.1007/s10329-023-01074-9.pdf

